|
森 哲 (京都大学大学院理学研究科)
マダガスカルを代表する爬虫類としてはカメレオンが有名である。爬虫類好きの人であれば、ヘラオヤモリ類もマダガスカルならではの動物として知っているだろう。しかし、それ以外のトカゲ類やヘビの仲間はそれほど印象的なものがいないせいか、一般にはあまり注目されていない。しかしながら、マダガスカルにおけるトカゲやヘビの多様性や固有種率は、キツネザル類に代表される哺乳類を凌ぐくらい、実は高いのである。
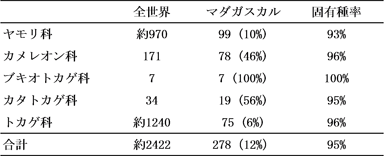
現生の爬虫類は4つのグループ(目)からなり、トカゲ類とヘビ類は合わせて有隣目を構成している。トカゲ類はさらに27のグループ(科)に細分される。このうちマダガスカルに分布するのはヤモリ科、カメレオン科、ブキオトカゲ科、トカゲ科、および、カタトカゲ科の5つである。種数が最も多いのはヤモリ科で現在約100種が確認されており、その約93%がマダガスカル固有種である(表1)。続いて種数が多いのはカメレオン科とトカゲ科で80種近くになり、固有種率は96%である。ブキオトカゲ科は広義のイグアナの仲間で、7種しか存在しないものの、マダガスカル固有の科である。カタトカゲ科はアフリカに分布する一見普通のトカゲだが、1種を除いてマダガスカル固有である。合計278種に及ぶトカゲ類全体で見ると、約95%がマダガスカルの固有種ということになる。
 |
 |
表1:
マダガスカルのトカゲ類の種数と固有種率。括弧内は全世界に対するマダガスカルの生息種数の割合を示す。種数に関しては、Glaw and Vences (2007)、Vitt and Caldwell (2009)を参考にした。 |
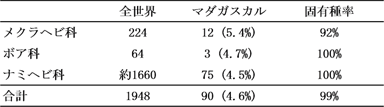
一方、ヘビ類は全世界に18科が存在する(最近では、さらに細分化して科の数を増やす傾向にあるが、ここでは従来の分類体系を採用しておく)。マダガスカルにはメクラヘビ科、ボア科、および、ナミヘビ科のみが分布する(ただし、コブラ科に属するセグロウミヘビが稀に沿岸で採集されることもある)。ナミヘビ科は非常に大きなグループで、全世界から約1660種が知られているが、その4.5%を占める75種がマダガスカルから報告されており、すべて固有種である(表2)。一方、いわゆる大蛇であるボア科は3種のみ生息し、これらもすべてマダガスカル固有である。メクラヘビ科は小型で地中棲の種が多いため、分布や分類学的な調査はマダガスカルで十分になされているとは言いがたいが、現在のところ12種が知られており、世界の熱帯域に広く分布する1種を除き、すべて固有種である。したがって、90種が生息するヘビ類に関しては、99%がマダガスカル固有の種となる。
表2:
マダガスカルのヘビ類の種数と固有種率。括弧内は全世界に対するマダガスカルの生息種数の割合を示す。種数に関しては、Glaw and Vences (2007)、Vitt and Caldwell (2009)を参考にした。
|
|
 |
マダガスカルにおけるヘビ類は、何が分布するかではなく、何が分布しないかという点においても際立った特徴がある。すなわち、いわゆる毒ヘビ類であるクサリヘビ科とコブラ科が生息しないのである。前者は日本ではマムシやハブに代表されるグループで、後者はその名の通りキングコブラなどを含み、日本でも琉球列島に小型種が2種分布している。
なぜマダガスカルにはこれらの毒ヘビ類が存在しないのか。クサリヘビ科やコブラ科は、1億6000万年くらい前にマダガスカルが大陸から分離したあとになって、大陸のどこかで新たに進化してきたと考えられている。そのため、これらの毒ヘビは海を越えてマダガスカルに渡ってこられず、現在も分布していないのだと、かつては漠然と考えられていた。しかしながら、最近の分子遺伝学的な研究によると、マダガスカルに現生するナミヘビ科は、2000万年~3200万年ほど前に大陸から海を越えて渡ってきた祖先に由来すると推測されている(表3)。では、どうしてナミヘビ科の仲間は海を超えてきたのに、コブラ類やクサリヘビ類はやってこなかったのか。何らかの生態的、生理的な要因のため、これらの毒ヘビ類は渡れなかったのか。あるいは、もしかしたら何度か渡ってきたものの、他のヘビ類との競争に負けて、現在は生き残っていないだけなのかもしれない。いずれにせよ、いわゆる「危険な毒ヘビ」が存在しないことは、マダガスカルの爬虫類相の大きな特徴の一つと言えるだろう。
 |
|
表3:
マダガスカルにおける現生ヘビ類の祖先の推定渡来時期、および、関係するヘビ類の推定分化時期。分化、侵入等の時期はNagy et al. (2003)、Noonanand Chippindale (2006)、Wuster et al. (2008)、Kelly et al. (2009)を参考にした。 |
1 Mimophis属を除くグループの共通祖先
2 Mimophis属の祖先
マダガスカルの爬虫類の多様性に関する調査は、島の東部を占める熱帯雨林におけるものが多く、西部の乾燥帯や中央高地はそれほど注目されていない。西部乾燥林帯に位置するアンピジュルア(Ampijoroa)では、京都大学を中心とする日本の大学とアンタナナリブ大学を含むマダガスカルの研究機関との共同調査が長年に渡って行われており、その成果の一つとして、爬虫類の多様性の高さが明らかになってきた。アンピジュルアの森はマダガスカル北西部のアンカラファンチカ(Ankarafantsika)国立公園の中心的な場所であり、熱帯落葉乾燥林帯に位置する。実際に調査を行っている範囲は2km×3km程度の狭い範囲だが、これまでに30種のトカゲ類と20種のヘビ類を確認している。この数は、マダガスカル全土に生息する種数のそれぞれ10.8%と22%に相当する。つまり、マダガスカルから知られているトカゲ類の10種に一つ、ヘビ類の5種に一つが、わずか6km²しかないアンピジュルアの森に生息していることになる。
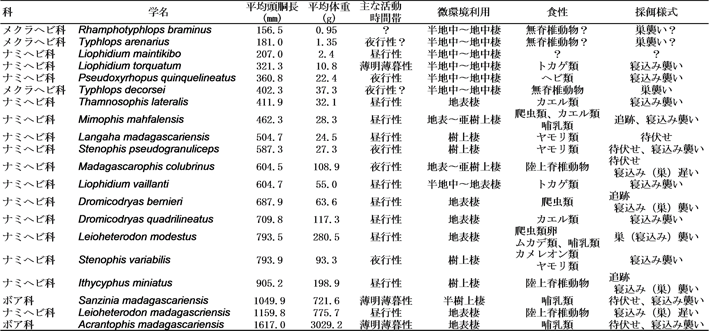
以下ではアンピジュルアのヘビ類の多様性について簡単に紹介する。ヘビ類は四肢の退化した細長い体形が特徴とされるが、相対的に大きな餌を丸呑みにするという摂食様式も、忘れてはならない独特の特徴である。ヘビ類はすべて肉食性であり、種ごとに餌として利用する動物群がほぼ決まっている。これに関連して、体の大きさ、頭の形、活動時間帯、微環境利用、採餌様式などが種ごとに異なっている。アンピジュルアの20種のヘビをこれらの視点から調査したところ、各種の頭胴長の平均は157~1617mm、体重の平均は0.95~3029gであり、この範囲内で途切れることなくサイズが多様化していた(表4)。過半数の種は主に昼行性であったが、3種は薄明薄暮型の活動性を示し、4種は主に夜行性と考えられた。地表棲または半地中棲の種が過半数を占め、樹上棲は5種程度であった。17種255個体のヘビから得られた690個の餌に基づくと、3種のヘビは哺乳類、鳥類、爬虫類、および、両棲類のすべての陸生脊椎動物群を餌とするジェネラリストであり、他の種は哺乳類専門食者、トカゲ類専門食者、カエル類専門食者などに分けられた。さらに、これらの食性、採餌行動の直接観察、および、餌動物との活動時間帯の比較などに基づき、各種の採餌様式を推測すると、待伏せ型、追跡型、寝込み襲い型、および、巣襲い型に分けられた。このように、アンピジュルアに生息する20種のヘビ類は、食性に関連した生態的、行動的要素において適応放散しており、これにより多くの種の共存が可能になっていると推測される。
マダガスカルのトカゲ類、ヘビ類は他の動物群と同様、非常に高い多様性や固有性を示す。また、その多様性は熱帯雨林だけでなく、アンカラファンチカのような落葉乾燥林においても実現しているのである。
表4:
アンピジュルア乾燥林に生息するヘビ類20種の大きさ、および、生態的・行動的諸要素。平均頭胴長の小さい順に記す。
参考文献
Glaw, F. and Vences, M. 2007. A Field Guide to the Amphibians and Reptiles of Madagascar. 3rd edition. Vences & Glaw Verlags GbR, Koln.
Kelly, C. M. R., Barker, N. P., Villet, M. H. and Broadley, D. G. 2009. Phylogeny, biogeography and classification of the snake superfamily Elapoidea: a rapid radiation in the late Eocene. Cladistics, 25: 38-63.
Nagy, Z. T., Joger, U., Wink, M., Glaw, F. and Vences M. 2003. Multiple colonization of Madagascar and Socotra by colubrid snakes: evidence from nuclear and mitochondrial gene phylogenies. Proc. R. Soc. Lond., B., 270: 2613-2621.
Noonan, B. P. and Chippindale, P. T. 2006. Dispersal and vicariance: the complex evolutionary history of boid snakes. Mol. Phylogenet. Evol., 40: 347-358.
Vitt, L. J. and Caldwell, J. P. 2009. Herpetology. An Introductory Biology of Amphibians and Reptiles. 3rd edition. Academic Press, Amsterdam.
Wuster, W., Peppin, L., Pook, C. E. and Walker, D. E. 2008. A nesting of vipers: phylogeny and historical biogeography of the Viperidae (Squamata: Serpentes). Mol. Phylogenet. Evol., 49: 445-459.
▲ページトップへ
|